Introduction
The ‘gut microbiota’ (GM) refers to the bacteria, viruses, fungi, and protozoans that colonize the gastrointestinal tract (GIT).1 The GM-host relationship is mutually beneficial, with the GM contributing to host homeostasis through digestion, vitamin synthesis, protection of the mucosal barrier, development of the immune system, and protection against pathogenic bacteria.2 Gut dysbiosis is an imbalance in the composition and function of the microbial communities in the gastrointestinal tract, leading to alterations in metabolic activity and immune response.3,4 The condition is typically characterized by a reduction in microbial diversity, a decrease in beneficial bacteria, and an overgrowth of harmful microorganisms.4 Over the last several decades, there has been a growing body of research that describes the impact of the gut microbiome on host physiology,2 with GM dysbiosis being linked to a variety of conditions including inflammatory bowel disease, cardiovascular diseases, cancer, respiratory diseases, diabetes, brain disorders, chronic kidney diseases, and liver diseases.5,6 Many studies point to the peripheral nervous system (PNS) as a way that the GM impacts peripheral organs, with the PNS transporting gut metabolites to distant tissues.7 However, more recently, researchers have been investigating the impact of the GM on the PNS, itself.
Peripheral neuropathies, affecting approximately 2% of the general public and 8% of older adults, include disorders of peripheral neurons and fibers, which manifest due to a wide variety of pathologies.8 The most common symptoms associated with peripheral neuropathy are numbness and paresthesias; however pain, weakness, and loss of deep tendon reflexes may also occur.8 This review aims to explore the current evidence regarding the relationship between the gut microbiome and peripheral nerve disorders with a focus on understanding the mechanisms through which microbial communities may contribute to the development and management of peripheral neuropathy and neuropathic pain. This analysis focused on sciatic nerve crush injury and chronic constriction injury (CCI) models, as these models are the most applicable to nerve injuries and compressive neuropathies. We seek to highlight the potential of gut microbiome-targeted therapies as novel interventions for these challenging conditions.
Methods
Literature Review Search
Using a modified PICO (problem, intervention, control, and outcome) method, a literature review was conducted to locate peer-reviewed articles in the PubMed and OVID/Medline databases. The key words used in two search components were (1) peripheral nerve injury, chronic constriction injury, CCI, nerve regeneration, nerve crush, neuropathic pain, nerve regeneration, peripheral nerve, nerve injury, innervation, or neuropathy, and (2) gut microbiota, gut microbiome, gastrointestinal microbiota, gastrointestinal microbiome, or dysbiosis.
Literature Inclusion
The titles and abstracts of articles identified through the search were reviewed by two reviewers. Any articles in the English language from all publication years through December 2024 were included for screening. Articles identified in the primary search were eligible for inclusion if they discussed the gut microbiome and peripheral nerve health. If the article was published in a supplemental section of a journal article as an abstract, it was excluded. If treatment groups had multiple variables or types of treatment compared to controls, the study was included if the GM intervention was assessed by itself as well. Additionally, articles were excluded if studies were not conducted in sciatic nerve crush injury or CCI animal models. The following information was extracted from the studies: species used, sample size, animal model used/nerve intervention, gut microbiota intervention and effect on GM composition, metabolic changes, key findings and conclusions, and the article’s proposed mechanism. The primary measures assessed were outcomes associated with GM interventions, categorized as motor function (sciatic function index [SFI]), sensory recovery (mechanical withdrawal threshold [MWT] and thermal withdrawal latency [TWL]), and histological changes.
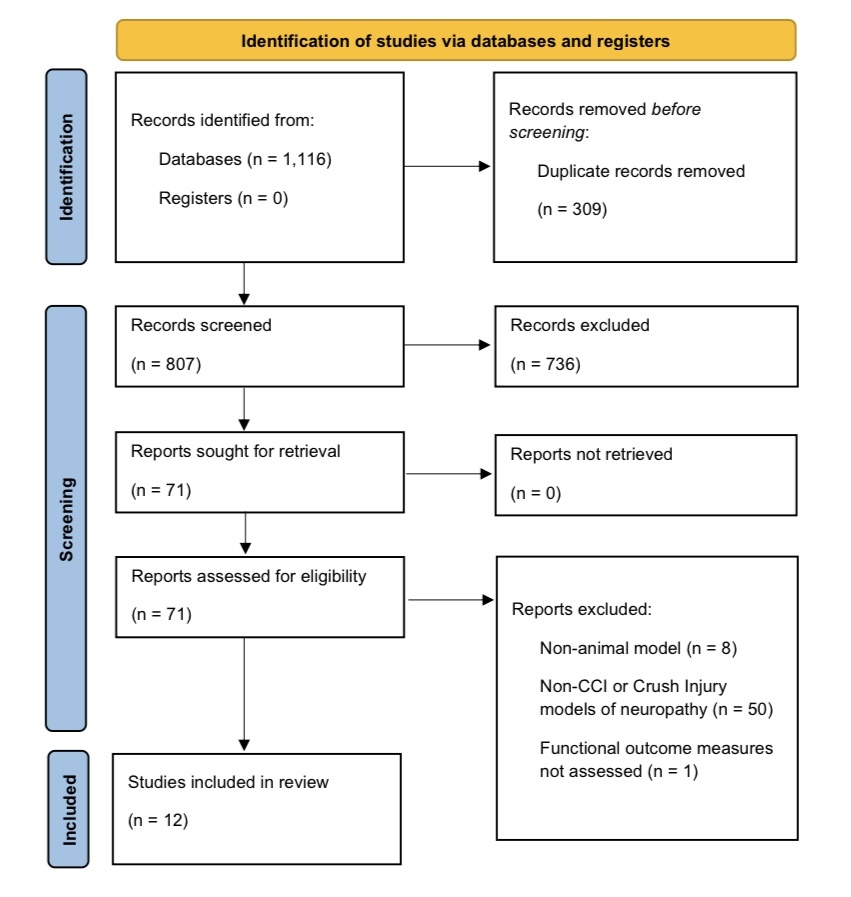
The primary search identified 1,116 articles, and 309 duplicates were removed. Of the remaining articles, 71 met criteria for inclusion for full analysis based on review of the abstracts. After full analysis of the articles, 12 articles were included in the final analysis (Supplementary Figure 1). Four studies evaluated sciatic nerve crush injury, and nine studies utilized a sciatic CCI model. Of the twelve studies, four evaluated motor function, eleven evaluated sensory recovery via MWT and TWL, and seven examined histological outcomes.
Risk of Bias
Investigators utilized the Systematic Review Center for Laboratory Animal Experimentation (SYRCLE) tool to evaluate all studies for bias (Supplementary Table 1).9 The allocation sequence was adequately generated and applied in 7 of the studies,10–16 while it was unclear in the other 5 studies17–21 if treatments were randomly assigned. All studies had similar baseline characteristics for the animal tested. It was documented in 1 study14 that the investigators were blinded to the treatment designation of the animals, while this was unclear in the other 11 studies.10–13,15–21 Outcome assessors were documented to be blinded in 5 of the 12 studies,10,12,14,17,21 while the other 7 studies11,13,15,16,18–20 did not disclose if assessors were blinded.
Gut Microbiota Interventions for Peripheral Nerve Recovery
Among the studies analyzed, five interventions targeting the gut microbiota were investigated in relation to peripheral nerve injury recovery. These interventions included probiotic (PBX) supplementation, antibiotic (ABX) treatment, intermittent fasting (IF), fecal microbiota transplantation (FMT), and naturopathic compounds such as Emodin and Danggui Sini Decoction (DGSD).
Probiotics
Peripheral nerve injuries have been shown to alter the composition of the gut microbiota.22 For this reason, researchers investigated whether treatment with PBX could improve recovery after PNI (Table 1). Although PBX treatment with Lactobacillus reuteri or Bifidobacterium did not result in significant changes in motor or sensory recovery after PNI,10 several studies showed improved sensory and motor recovery with treatment of a larger variety of bacterial strains in commonly used PBX cocktails.11,16,17 Wang et al. reported improvements in toe spreading, stride length, and weight of the gastrocnemius muscle with treatment of PBX cocktail compared to control CCI mice, suggesting that PBX treatment improves motor recovery following sciatic nerve CCI.11 Additionally, PBX treatment improved motor recovery, measured by SFI, compared to control.16 In terms of sensory recovery, PBX improved mechanical and thermal hyperalgesia, measured by MWT and TWL, after sciatic nerve CCI.11,17 Histologically, PBX treatment improved the gastrocnemius muscle fiber size, which was significantly reduced after sciatic CCI.11 These results indicate that probiotic treatment may improve recovery of both sensory and motor function following peripheral nerve injury.
Antibiotics
Researchers aimed to investigate whether eradication of the GM with ABX treatment would impact recovery after PNI (Table 2). Studies investigating the impact of ABX treatment on recovery following PNI showed that ABX treatment prevented thermal and mechanical hyperalgesia after sciatic CCI.12,18,21 However, Ma et al. found that fecal microbiota transplantation from a control mouse reversed this effect. Additionally, Zhou et al. found that mice treated with oral administration of SCFAs after ABX+CCI displayed thermal and mechanical hyperalgesia, suggesting that the SCFAs produced by the gut microbiota are required for the development of neuropathic pain.18 In the sciatic crush injury model, pre- or post-injury ABX treatment impaired motor recovery, as measured by Sciatic Function Index (SFI).16 However, this impairment in motor recovery was prevented by the co-treatment of PBX, indicating that PBX treatment can rescue the functional deficits seen with ABX treatment.16 Additionally, ABX treatment suppressed the increase in activation of microglial cells (Iba1) and astrocytes (GFAP) in the spinal cord, normally observed after CCI.21 These results suggest that although ABX treatment negatively impacts recovery after PNI, these negative effects can be reversed with reconstitution of the GM through co-treatment with PBX or FMT.
Intermittent Fasting
Intermittent fasting (IF) involves the time-based restriction of food intake that improves richness and alters the composition of the gut microbiota.23 Additionally, IF has previously been shown to promote metabolic and signaling pathways that increase synaptic plasticity and neurogenesis, which are vital for axonal regeneration.24–27 In the articles reviewed, researchers investigated the impact of IF on recovery after PNI, as well as the differential metabolites observed after IF that could contribute to these effects (Table 3). Intermittent fasting impacted the serum levels of gut microbiota-derived metabolites, increasing indole-3-propionate (IPA) and decreasing imidazole propionate (ImP). In both studies, administration of IPA and ImP, respectively, were also evaluated individually for their effect on recovery after PNI, outside of IF treatment. ImP is a histidine-derived gut metabolite, which has been correlated with a variety of pathologies, including diabetes and cardiometabolic diseases in previous studies.28–30 Tang et al. found that ImP treatment reduced sensory and motor recovery after sciatic crush injury.19 Histologically, ImP treatment inhibited Schwann cell migration, and both myelin thickness and density of axons were decreased compared to the control CCI group.19 Schwann cells make up the peripheral nerve myelin sheath, and their migration to the injury site are vital to repair after peripheral nerve injury.31
IPA is a Tryptophan-derived indole compound produced by specific microbial taxa, which has been shown to protect against oxidative stress, attenuate inflammation, and improve the intestinal barrier integrity by inducing the expression of tight junction proteins such as ZO-1 and Occludin.32,33 IPA has also been studied for its potentially positive effects in cardiovascular disease and Alzheimer’s disease.32,33 Serger et al. found that IPA improved sensory recovery after sciatic crush injury through neutrophil chemotaxis.14
Fecal Microbiota Transplantation
Fecal microbiota transplantation (FMT) involves transplanting stool from a donor to a recipient and can be useful tool in gut microbiota research. For example, FMT was utilized by Serger et al. to identify if the microbial changes induced by IF would promote sciatic nerve regeneration.14 Stool from the IF group was transplanted to the control sciatic nerve crush group through FMT, and this resulted in an increase in regenerated nerve fibers past the sciatic nerve crush site in recipients of the FMT.14
FMT from a healthy control can also be used to normalize the GM composition after PNI (Table 4). FMT from control young mice to aged sciatic nerve crush injury mice resulted in improvement in mechanical and thermal hyperalgesia, compared to control mice.20 Mice that underwent FMT displayed increased expression of anti-inflammatory IL-10 in the damaged sciatic nerve.20 Additionally, mice in the FMT group had increased expression of Claudin-1, indicating a tighter mucosal barrier.20
Svacina et al. proposed short-chain fatty acids (SCFAs), including butyrate, acetate, and propionate, as a potential mechanism of the gut microbiota for modulating the inflammatory response and improving recovery after PNI.20 There was an increased abundance of serum SCFA in the mice that underwent FMT, compared to the control group.20 SCFAs are essential for maintaining the gut barrier integrity and modulating immune responses, including reducing pro-inflammatory cytokines and promoting regulatory T-cell activity.34 Adding to the evidence of the importance of SCFAs in nerve regeneration, researchers have investigated the impact of monocarboxylate transporter 1 (MCT1) deficiency on peripheral nerve regeneration after sciatic nerve crush injury.35 MCT1 is localized within the peripheral nervous system in perineurial cells, dorsal root ganglion neurons, and Schwann cells and is responsible for the transport of monocarboxylates, including SCFAs, across cell membranes.35 Additionally, MCT1 is found in macrophages and is vital for normal macrophage metabolism.36 Researchers found that selective ablation of MCT1 in macrophages impaired nerve regeneration, while MCT1 overexpression in macrophages accelerated peripheral nerve regeneration.36 Additionally, in MCT1 deficient mice, nerve regeneration was delayed following sciatic nerve crush injury.35 These studies demonstrate the importance of short chain fatty acids, which are produced by specific bacterial taxa within the gut.36
Naturopathic Compounds
A variety of other dietary changes aimed at modulation of the GM were also studied, including Danggui Sini Decoction (DGSD) and Emodin (Table 5).13,15 Emodin (6-methyl-1,3,8-trihydroxyanthraquinone) is an anthraquinone that can be derived from several commonly used Chinese herbs including Rheum palmatum, Polygonum cuspidatum and Polygonum multiflorum.37 Wang et al. observed improved motor and sensory recovery after sciatic nerve CCI with the treatment of Emodin, compared to an isolated CCI group.15 Emodin reduced the expression of pro-inflammatory cytokines, including IL-6, IL-1B, and TNF-α in both spinal cord and intestines.15 Additionally, Emodin increased the expression of the tight junction proteins ZO-1 and Occludin, indicating a function in maintaining intestinal barrier integrity.15
DGSD, an herbal mixture utilized in traditional Chinese medicine, normalized the gut microbiota profile after sciatic nerve CCI and improved thermal hyperalgesia after sciatic nerve CCI, although sensory function was not returned to baseline.13 DGSD reduced expression of pro-inflammatory cytokines, including IL-1α, SP, CCL5, PGE2, IL-6, COX2, MMP9, and increased expression of the anti-inflammatory cytokine IL-10 in the blood and sciatic nerve.13 Additionally, treatment with DGSD inhibited nNOS and NF-κB phosphorylation, which drive inflammation and oxidative damage.38,39 Histologically, DGSD decreased inflammatory cell infiltration compared to isolated CCI, although this was not returned to baseline.13 These results suggest that Emodin and DGSD may improve recovery and decrease expression of pro-inflammatory cytokines after PNI.
Summary
The objective of this review was to examine the existing evidence on the relationship between gut microbiota and peripheral nerve health, with a particular focus on its potential relevance to peripheral nerve injuries and compressive neuropathies. The reviewed studies demonstrate that alterations in the gut microbiota, whether induced by antibiotics, probiotics, intermittent fasting, or dietary interventions, impact functional recovery, neuropathic pain, and inflammatory processes. Each of the therapies reviewed was considered safe in the evaluated studies. None of the studies included in this review reported negative or damaging effects on nerve tissue from the gut microbiota intervention. However, variability in probiotic formulations and treatment durations highlights the need for standardized clinical studies. Previous literature has described quantitative differences between rodent and human gut microbiota, but qualitatively, the GM of both species were relatively similar.40 While animal studies provide valuable insights into mechanisms of action, their relevance to human PNI remains uncertain without a clearer understanding of how these microbial interactions manifest in humans. Future research should focus on identifying the specific microbial taxa responsible for neuroregenerative effects and developing standardized protocols that account for interspecies differences. By improving our understanding of these translational challenges, microbiota-based therapies for PNI can be more effectively adapted for human patients.
Acknowledgements
None
Author Contributions Statement
All authors have read and approved the final submitted manuscript. The following is the author contribution: MM, BR, JAVN: conceptualization; MM, SR, GH, LS: article screening and bias assessment; MM, BR, JAVN, SR, GH, LS: manuscript writing and editing.
Conflict of Interest Disclosures
-
Julia A.V. Nuelle receives research support from Axogen; receives research support from Musculoskeletal Transplant Foundation; is an paid consultant for Arthrex, Inc.; is a paid speaker for Trimed; is on the editorial or governing board and is a board or committee member for Arthroscopy; is a board or committee member of Society of Military Orthopaedic Surgeons; is a board or committee member of American Society for Surgery of the Hand; is a board or committee member for American Orthopaedic Association; and is a board or committee member of American Academy of Orthopaedic Surgeons.
-
Madeline K. Mueller, Sarah R. Rousseau, Gillian M. Hark, Lauren R. Shaffer, and Bryce F. Rizvanović declare no conflicts of interest.